

Световая фаза фотосинтеза
В световой фазе фотосинтеза происходит синтез АТФ и НАДФ·H2 за счет лучистой энергии. Это происходит на тилакоидах хлоропластов, где пигменты и ферменты образуют сложные комплексы для функционирования электрохимических цепей, по которым передаются электроны и отчасти протоны водорода.
Электроны в конечном итоге оказываются у кофермента НАДФ, который, заряжаясь отрицательно, притягивает к себе часть протонов и превращается в НАДФ·H2. Также накопление протонов по одну сторону тилакоидной мембраны и электронов по другую создает электрохимический градиент, потенциал которого используется ферментом АТФ-синтетазой для синтеза АТФ из АДФ и фосфорной кислоты.
Главными пигментами фотосинтеза являются различные хлорофиллы. Их молекулы улавливают излучение определенных, отчасти разных спектров света. При этом некоторые электроны молекул хлорофилла переходят на более высокий энергетический уровень. Это неустойчивое состояние, и по-идее электроны путем того же излучения должны отдать в пространство полученную из вне энергию и вернуться на прежний уровень. Однако в фотосинтезирующих клетках возбужденные электроны захватываются акцепторами и с постепенным уменьшением своей энергии передаются по цепи переносчиков.
На мембранах тилакоидов существуют два типа фотосистем, испускающих электроны при действия света. Фотосистемы представляют собой сложный комплекс большей частью хлорофильных пигментов с реакционным центром, от которого и отрываются электроны. В фотосистеме солнечный свет ловит множество молекул, но вся энергия собирается в реакционном центре.
Электроны фотосистемы I, пройдя по цепи переносчиков, восстанавливают НАДФ.
Энергия электронов, оторвавшихся от фотосистемы II, используется для синтеза АТФ. А сами электроны фотосистемы II заполняют электронные дырки фотосистемы I.
Дырки второй фотосистемы заполняются электронами, образующимися в результате фотолиза воды. Фотолиз также происходит при участии света и заключается в разложении H2O на протоны, электроны и кислород. Именно в результате фотолиза воды образуется свободный кислород. Протоны участвуют в создании электрохимического градиента и восстановлении НАДФ. Электроны получает хлорофилл фотосистемы II.
Примерное суммарное уравнение световой фазы фотосинтеза:
H2O + НАДФ + 2АДФ + 2Ф → ½O2 + НАДФ · H2 + 2АТФ
Циклический транспорт электронов
Выше описана так называемый нецикличная световая фаза фотосинтеза. Есть еще циклический транспорт электронов, когда восстановления НАДФ не происходит. При этом электроны от фотосистемы I уходят на цепь переносчиков, где идет синтез АТФ. То есть эта электрон-транспортная цепь получает электроны из фотосистемы I, а не II. Первая фотосистема как бы реализует цикл: в нее возвращаются ей же испускаемые электроны. По дороге они тратят часть своей энергии на синтез АТФ.
Фотофосфорилирование и окислительное фосфорилирование
Световую фазу фотосинтеза можно сравнить с этапом клеточного дыхания — окислительным фосфорилированием, которое протекает на кристах митохондрий. Там тоже происходит синтез АТФ за счет передачи электронов и протонов по цепи переносчиков. Однако в случае фотосинтеза энергия запасается в АТФ не для нужд клетки, а в основном для потребностей темновой фазы фотосинтеза. И если при дыхании первоначальным источником энергии служат органические вещества, то при фотосинтезе – солнечный свет. Синтез АТФ при фотосинтезе называется фотофосфорилированием, а не окислительным фосфорилированием.
Световая фаза
Световая фаза фотосинтеза начинается со сбора световой энергии в светособирающих комплексах молекул хлорофилла и передачи ее в реакционные центры этих комплексов. Молекулы хлорофилла расположены в клетках растений не хаотически.
Они соединяются друг с другом особыми энергетическими связями. Поглощая квант света, молекула хлорофилла получает частицу энергии, которую она передает по энергетическим связям к молекуле, называемой молекулой-ловушкой или реакционным центром.
На 200-400 молекул хлорофилла приходится один реакционный центр.
Каждая молекула хлорофилла может получить фотон света достаточно редко. Но чтобы процессы в растительной клетке не прерывались, фотоны собираются с некоторого участка листа в один реакционный центр.
В него световая энергия будет поступать намного чаще, чем из отдельных молекул хлорофилла. И химические реакции здесь будут идти практически непрерывно. Получив световую энергию, реакционные центры проводят фотолиз воды, которая в достаточном количестве находится в тканях клетки.
Формула фотолиза воды может быть представлена следующим образом;
2Н20 + Qсвета -> 4Н+ + 4е— + 02
Вода под воздействием световой энергии распадается на следующие составляющие:
- Протоны водорода (Н+);
- Электроны водорода (е—);
- Кислород (О2).
Но это еще не конечные продукты световой фазы фотосинтеза. Протоны идут на восстановление НАДФ до НАДФН. НАДФ (никотинамидадениндинуклеотидфосфат) – соединение, присутствующее во всех растительных и животных клетках и принимающее участие в фотосинтетических процессах.









Это фермент, играющий роль катализатора. Он принимает на себя протоны водорода, превращаясь в НАДФН, а затем отдает их в процессе дальнейших химических реакций. Восстановленный НАДФН и является источником водорода, который используется на темновой стадии фотосинтеза для получения глюкозы из углекислого газа.
Электроны при фотолизе воды восстанавливают АДФ до АТФ. АТФ (аденозинтрифосфат) – органическая молекула, присутствующая во всех живых клетках.
Она является источником энергии в различных химических реакциях. В них молекула АТФ переходит в АДФ (аденозиндифосфат), выделяя огромное количество энергии. АТФ – является один из конечных продуктов световой фазы фотосинтеза у растений – энергетическая составляющая, необходимая для прохождения химических реакций темновой фазы.
При фотолизе воды образуется кислород. Так как он не принимает участия в дальнейших химических процессах, растение выделяет его в окружающую среду. Итак, конечными продуктами световой фазы фотосинтеза являются:
- НАДФН;
- АТФ;
- Молекулярный кислород.
Световую фазу фотосинтеза можно представить следующей формулой:
Н20 + Qсвета -> НАДФН + АТФ + 02
Хлоропласты .
Хлоропласты – это структуры, в которых происходят фотосинтетические процессы, приводящие в конечном итоге к связыванию углекислоты, к выделению кислорода и синтезу сахаров.Структуры удлиненной формы с шириной 2-4 мкм и протяженностью 5-10 мкм. У зеленых водорослей встречаются гигантские хлоропласты (хроматофоры), достигающие длины 50 мкм.
у зеленых водорослей может быть по одному хлоропласту на клетку. Обычно на клетку высших растений приходится в среднем 10-30 хлоропластов. Встречаются клетки с огромным количеством хлоропластов. Например, в гигантских клетках палисадной ткани махорки обнаружено около 1000 хлоропластов.
Хлоропласты представляют собой структуры, ограниченные двумя мембранами – внутренней и внешней. Внешняя мембрана, как и внутренняя, имеет толщину около 7 мкм, они отделены друг от друга межмембранным пространством около 20-30 нм. Внутренняя мембрана хлоропластов отделяет строму пластиды, аналогичную матриксу митохондрий. В строме зрелого хлоропласта высших растений видны два типа внутренних мембран. Это – мембраны, образующие плоские, протяженные ламеллы стромы, и мембраны тилакоидов, плоских дисковидных вакуолей или мешков.
Ламеллы стромы (толщиной около 20 мкм) представляют собой плоские полые мешки или же имеют вид сети из разветвленных и связанных друг с другом каналов, располагающихся в одной плоскости. Обычно ламеллы стромы внутри хлоропласта лежат параллельно друг другу и не образуют связей между собой.
Кроме мембран стромы в хлоропластах обнаруживаются мембранные тилакоиды. Это плоские замкнутые мембранные мешки, имеющие форму диска. Величина межмембранного пространства у них также около 20-30 нм. Такие тилакоиды образуют стопки наподобие столбика монет, называемые гранами.
Число тилакоидов на одну грану очень варьирует: от нескольких штук до 50 и более. Размер таких стопок может достигать 0,5 мкм, поэтому граны видны в некоторых объектах в световом микроскопе. Количество гран в хлоропластах высших растений может достигать 40-60. Тилакоиды в гране сближены друг с другом так, что внешние слои их мембран тесно соединяются; в месте соединения мембран тилакоидов образуется плотный слой толщиной около 2 нм. В состав граны кроме замкнутых камер тилакоидов обычно входят и участки ламелл, которые в местах контакта их мембран с мембранами тилакоидов тоже образуют плотные 2-нм слои. Ламеллы стромы, таким образом, как бы связывают между собой отдельные граны хлоропласта. Однако полости камер тилакоидов всезда замкнуты и не переходят в камеры межмембранного пространства ламелл стромы. Ламеллы стромы и мембраны тилакоидов образуются путем отделения от внутренней мембраны при начальных этапах развития пластид.
В матриксе (строме) хлоропластов обнаруживаются молекулы ДНК, рибосомы; там же происходит первичное отложение запасного полисахарида, крахмала, в виде крахмальных зерен.
Характерным для хлоропластов является наличие в них пигментов, хлорофиллов, которые и придают окраску зеленым растениям. При помощи хлорофилла зеленые растения поглощают энергию солнечного света и превращают ее в химическую.
В хлоропластах содержатся различные пигменты. В зависимости от вида растений это:
хлорофилл:
— хлорофилл А (сине-зеленый) — 70 % (у высших растений и зеленых водорослей);
— хлорофилл В (желто-зеленый) — 30 % (там же);
— хлорофилл С, D и E встречается реже — у других групп водорослей;
каротиноиды:
— оранжево-красные каротины (углеводороды);
— желтые (реже красные) ксантофиллы (окисленные каротины). Благодаря ксантофиллу фикоксантину хлоропласты бурых водорослей (феопласты) окрашены в коричневый цвет;
фикобилипротеиды, содержащиеся в родопластах (хлоропластах красных и сине-зеленых водорослей):
— голубой фикоцианин;
— красный фикоэритрин.
Клеточная стенка растительной клетки: общие сведения
Клеточная стенка (нередко в качестве синонима термина «клеточная
стенка» в учебной и научной литературе используется термин
«клеточная оболочка».) у растений — это структурное образование,
располагающееся по периферии клетки, за пределами
плазмалеммы
, придающее клетке прочность, сохраняющее ее форму и защищающее
протопласт
.










Клеточная стенка растений противостоит высокому осмотическому давлению
большой центральной
вакуоли
и препятствует разрыву клетки. Кроме того, совокупность прочных клеточных
стенок выполняет роль своеобразного внешнего скелета, поддерживающего форму
растения и придающего ему механическую прочность. Клеточная стенка, обладая
большой прочностью, в то же время способна к росту, и прежде всего к росту
растяжением. Эти два в известной степени противоположных требования
удовлетворяются за счет особенностей ее строения и химического состава.
Клеточная стенка, как правило, прозрачна и хорошо пропускает солнечный
свет. Через нее легко проникают вода и низкомолекулярные вещества, но для
высокомолекулярных веществ она полностью или частично непроницаема. У
многоклеточных организмов стенки соседних клеток скреплены между собой
пектиновыми веществами, образующими срединную пластинку.
При специальной обработке растительных тканей некоторыми веществами
(крепкие щелочи, азотная кислота) стенки соседних клеток разъединяются в
результате разрушения срединной пластинки. Этот процесс называется
мацерацией
. Естественная мацерация происходит у перезрелых плодов груши, дыни,
персика и др.
В результате тургорного давления стенки соседних клеток в углах могут
округляться и между ними образуются межклетники.
Стенка клетки представляет собой продукт жизнедеятельности ее
протопласта
. Поэтому стенка может расти, только находясь в контакте с протопластом.
Однако при отмирании протопласта стенка сохраняется и мертвая клетка может
продолжать выполнять функции проведения воды или играть роль механической
опоры.
Основу клеточной стенки составляют высокополимерные углеводы: молекулы
целлюлозы (клетчатки)
, собранные в сложные пучки — фибриллы, образующие каркас, погруженный в
основу (матрикс), состоящий из
гемицеллюлоз
,
пектинов
и
гликопротеидов
(
рис. 21
). Молекулы
целлюлозы
состоят из большого числа линейно расположенных мономеров — остатков
глюкозы
. Целлюлоза очень стойка, не растворяется в разбавленных кислотах и даже в
концентрированных щелочах. Эластичный целлюлозный скелет придает клеточной
оболочке механическую прочность. Первоначально число микрофибрилл,
образованных молекулами целлюлозы, в клеточной стенке относительно
невелико, но с возрастом оно увеличивается и клетка теряет способность к
растяжению.
Гемицеллюлозы
отличаются от
целлюлозы
составом мономеров и разветвленным их расположением в молекулах. Являясь
одним из компонентов пластичного матрикса,
гемицеллюлозы
придают клеточной стенке дополнительную прочность, но почти не
препятствуют ее росту.
Гемицеллюлозы
могут быть и запасными веществами, так как легко гидролизуются. Кроме
гемицеллюлоз в матрикс, а также в срединную пластинку входят пектиновые
вещества, или
пектины
, и
полисахариды
, образованные мономерами —
уроновыми кислотами
. Эти вещества скрепляют, склеивают оболочки соседних клеток. Молекулы
гемицеллюлоз, пектина и гликопротеидов соединяют целлюлозные
микрофибриллы.
Помимо
полисахаридов
, в матриксе стенок многих клеток часто обнаруживаются неуглеводные
компоненты. Наиболее обычен из них
лигнин
— полимерное вещество полифенольной природы. Содержание его в стенках
некоторых видов клеток может достигать 30%.
Лигнин
откладывается при завершении роста стенки. Процесс отложения лигнина
получил название
одревеснения, или лигнификации
. Стенка, пропитанная лигнином, очень прочна и тверда. Лигнифицируются
чаще всего оболочки клеток, подвергающихся механическим нагрузкам.
Стенки некоторых типов клеток могут включать слои
липидов
:
воска
,
кутина
и
суберина
.
Кутин
и
воск
обычно покрывают наружные стенки клеток эпидермы. Слой кутина создает на
поверхности растения водо- и воздухонепроницаемый слой
кутикулы
. Суберин пропитывает стенки. Он непроницаем для воды и газов, поэтому
такая суберинизированная, или опробковевшая, клетка быстро отмирает.
Значение фотосинтеза в природе
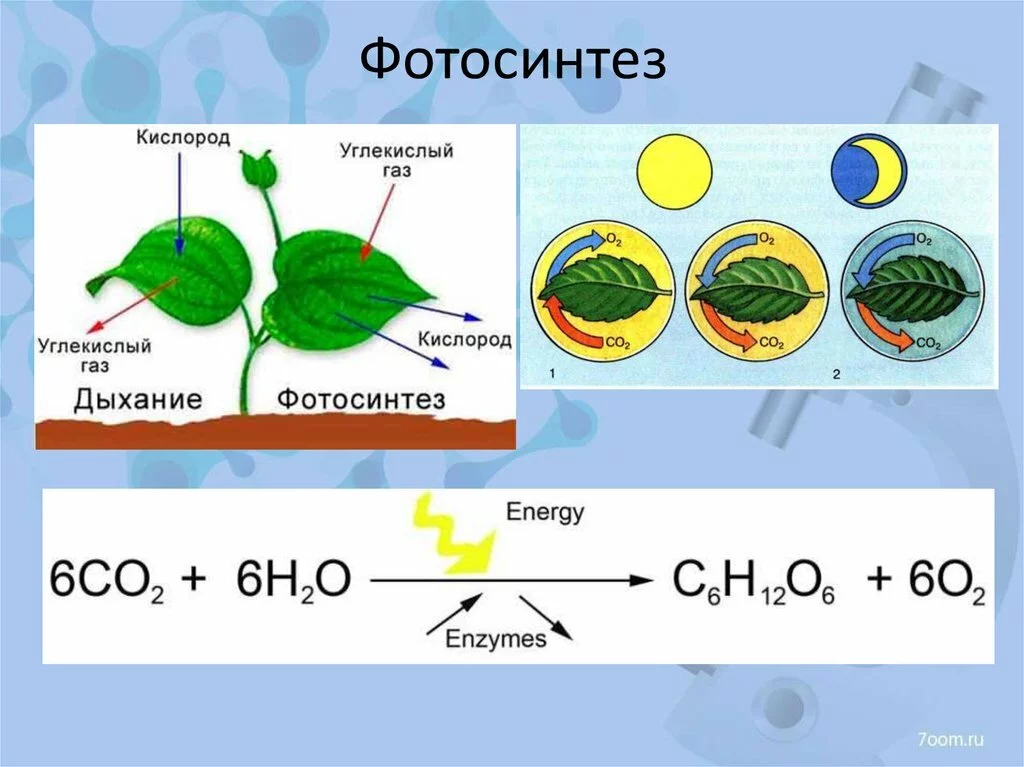
Биохимический процесс фотосинтеза использует энергию солнечного света для преобразования воды и углекислого газа в кислород и глюкозу. Глюкоза используется в качестве строительных блоков в растениях для роста тканей. Таким образом, фотосинтез — это способ, благодаря которому формируются корни, стебли, листья, цветы и плоды. Без процесса фотосинтеза растения не смогут расти или размножаться.
Продуценты
Из-за фотосинтетической способности, растения известны как продуценты и служат основой почти каждой пищевой цепи на Земле. (Водоросли являются эквивалентом растений в ). Вся пища, которую мы едим, происходит от организмов, являющихся фотосинтетиками. Мы питаемся этими растениями напрямую или едим животных, таких как коровы или свиньи, которые потребляют растительную пищу.
Основа пищевой цепи
Внутри водных систем, растения и водоросли также составляют основу пищевой цепи. Водоросли служат пищей для беспозвоночных, которые, в свою очередь, выступают источником питания для более крупных организмов. Без фотосинтеза в водной среде жизнь была бы невозможна.
Удаление углекислого газа
Фотосинтез превращает углекислый газ в кислород. Во время фотосинтеза углекислый газ из атмосферы поступает в растение, а затем выделяется в виде кислорода. В сегодняшнем мире, где уровни двуокиси углерода растут ужасающими темпами, любой процесс, который устраняет углекислый газ из атмосферы, является экологически важным.
Круговорот питательных веществ
Растения и другие фотосинтезирующие организмы играют жизненно важную роль в круговороте питательных веществ. Азот в воздухе фиксируется в растительных тканях и становится доступным для создания белков. Микроэлементы, находящиеся в почве, также могут быть включены в растительную ткань и стать доступными для травоядных животных, дальше по пищевой цепи.
Фотосинтетическая зависимость
Фотосинтез зависит от интенсивности и качества света. На экваторе, где солнечный свет обилен весь год и вода не является ограничивающим фактором, растения имеют высокие темпы роста, и могут стать довольно большими. И наоборот, фотосинтез в более глубоких частях океана встречается реже, поскольку свет не проникает в эти слои, и в результате эта экосистема оказывается более бесплодной.










Г) Эндоплазматическая сеть (ЭПС) — Студопедия
Вся внутренняя зона цитоплазмы заполнена многочисленными мелкими каналами и полостями, стенки которых представляют собой мембраны, сходные по своей структуре с плазматической мембраной.
Эти каналы ветвятся, соединяются друг с другом и образуют сеть, получившую название эндоплазматической сети. Эндоплазматическая сеть неоднородна по своему строению.
Известны два ее типа – гранулярная и гладкая.
На мембранах каналов и полостей гранулярной сети располагается множество мелких округлых телец – рибосом, которые придают мембранам шероховатый вид. Мембраны гладкой эндоплазматической сети не несут рибосом на своей поверхности. ЭПС выполняет много разнообразных функций.
Основная функция гранулярной эндоплазматической сети – участие в синтезе белка, который осуществляется в рибосомах. На мембранах гладкой эндоплазматической сети происходит синтез липидов и углеводов.
Все эти продукты синтеза накапливаются в каналах и полостях, а затем транспортируются к различным органоидам клетки, где потребляются или накапливаются в цитоплазме в качестве клеточных включений. ЭПС связывает между собой основные органоиды клетки(рис. 2.13).
Рис. 2.13. Строение эндоплазматической сети (ЭПС) или ретикулума
Д) Аппарат Гольджи
Строение этого органоида сходно в клетках растительных и животных организмов, несмотря на разнообразие его формы. Выполняет много важных функций.
Одномембранный органоид. Представляет собой стопки уплощенных «цистерн» с расширенными краями, с которыми связана система мелких одномембранных пузырьков (пузырьки Гольджи).
Пузырьки Гольджи в основном сконцентрированы на стороне, примыкающей к ЭПС, и по периферии стопок.
Полагают, что они переносят в аппарат Гольджи белки и липиды, молекулы которых, передвигаясь из цистерны в цистерну, подвергаются химической модификации.
Все эти вещества сначала накапливаются, химически усложняются, а затем в виде крупных и мелких пузырьков поступают в цитоплазму и либо используются в самой клетке в процессе ее жизнедеятельности, либо выводятся из нее и используются в организме (рис. 2.14-2.15).
Рис. 2.14. Строение аппарата Гольджи
Функции:
– накопление белков, липидов, углеводов;
– модификация и упаковка в мембранные пузырьки (везикулы) поступивших органических веществ; секреция белков, липидов, углеводов;
– место образования лизосом.
– секреторная функция, поэтому аппарат Гольджи хорошо развит в секреторных клетках.
Рис. 2.15. Комплекс Гольджи










Е) Лизосомы
Представляют собой небольшие округлые тельца. Внутри лизосомы находятся ферменты, расщепляющие белки, жиры, углеводы, нуклеиновые кислоты. К пищевой частице, поступившей в цитоплазму, подходят лизосомы, сливаются с ней, и образуется одна пищеварительная вакуоль, внутри которой находится пищевая частица, окруженная ферментами лизосом.
Одномембранные органоиды. Представляют собой мелкие пузырьки (диаметр от 0,5 до 2 мкм), содержащие набор гидролитических ферментов.
Ферменты синтезируются на шероховатой ЭПС, перемещаются в аппарат Гольджи, где происходит их модификация и упаковка в мембранные пузырьки лизосом. Лизосома может содержать от 20 до 60 различных видов гидролитических ферментов.
Расщепление веществ с помощью ферментов называют лизисом. Различают первичные и вторичные лизосомы. Первичными называются лизосомы, отпочковавшиеся от аппарата Гольджи.
Вторичными называются лизосомы, образовавшиеся в результате слияния первичных лизосом с эндоцитозными вакуолями. В этом случае в них происходит переваривание веществ, поступивших в клетку путем фагоцитоза или пиноцитоза, поэтому их можно назвать пищеварительными вакуолями.
Функции лизосом:
– переваривание захваченных клеткой при эндоцитозе веществ или частиц (бактерий, других клеток),
– аутофагия — уничтожение ненужных клетке структур, например, во время замены старых органоидов новыми, или переваривание белков и других веществ, произведенных внутри самой клетки,
– автолиз — самопереваривание клетки, приводящее к ее гибели (иногда этот процесс не является патологическим, а сопровождает развитие организма или дифференцировку некоторых специализированных клеток) (рис. 2.16-2.17).
Пример: При превращении головастика в лягушку, лизосомы, находящиеся в клетках хвоста, переваривают его: хвост исчезает, а образовавшиеся во время этого процесса вещества всасываются и используются другими клетками тела.
Рис. 2.16. Образование лизосом
Рис. 2.17. Функционирование лизосом
Митохондрии
Митохондрии — органеллы, характерные для большинства клеток растений. Имеют изменчивую форму палочек, зёрнышек, нитей. Открыты в 1894 году Р. Альтманом с помощью светового микроскопа, а внутреннее строение было изучено позднее с помощью электронного.
Строение митохондрии
Митохондрии имеют двухмембранное строение. Внешняя мембрана гладкая, внутренняя образует различной формы выросты — трубочки в растительных клетках. Пространство внутри митохондрии заполнено полужидким содержимым (матриксом), куда входят ферменты, белки, липиды, соли кальция и магния, витамины, а также РНК, ДНК и рибосомы. Ферментативный комплекс митохондрий ускоряет работу сложного и взаимосвязанного механизма биохимических реакций, в результате которых образуется АТФ. В этих органеллах осуществляется обеспечение клеток энергией — преобразование энергии химических связей питательных веществ в макроэргиеские связи АТФ в процессе клеточного дыхания. Именно в митохондриях происходит ферментативное расщепление углеводов, жирных кислот, аминокислот с освобождением энергии и последующим превращением её в энергию АТФ. Накопленная энергия расходуется на ростовые процессы, на новые синтезы и т. д. Митохондрии размножаются делением и живут около 10 дней, после чего подвергаются разрушению.
Пигменты пластид
В структуру клеточных органоидов входят три группы пигментов:
- хролофилл – магний-порфириновые белковые комплексы хромопротеидов, придающие листьям, стволу зеленую окраску;
- каротиноид – красящий пигмент, схожий с ретинолом (витамин А), в зависимости от концентрации обретают оранжевую или красноватую окраску;
- ксантофилл по сути – окисленный каротин, содержится вместе с р-каротином, имеет такие же функции;
- фикобилинпротеиды по компонентной структуре схожи с желчными пигметно-белковыми соединениями. К ним относятся: синие фикоцианины, придающие окраску плодам; красно-бордовые фикоэритрины.
Доказательства
Относительно недавно гипотеза о прокариотическом происхождении этих элементов была не слишком популярна в научном сообществе, многие считали ее «измышлениями дилетантов». Но после того как был проведен углубленный анализ нуклеотидных последовательностей в ДНК хлоропластов, это предположение получило блестящее подтверждение. Выяснилось, что эти структуры чрезвычайно схожи, даже родственны, ДНК бактериальных клеток. Так, аналогичная последовательность была найдена у свободноживущих цианобактерий. В частности, оказались чрезвычайно схожи гены АТФ-синтезирующего комплекса, а также в «аппаратах» транскрипции и трансляции.










Промоторы, которые определяют начало считывания генетической информации с ДНК, а также терминальные нуклеотидные последовательности, которые отвечают за ее прекращение, также организованы по образу и подобию бактериальных. Разумеется, миллиарды лет эволюционных преобразований смогли внести множество изменений в хлоропласт, но последовательности в хлоропластных генах остались абсолютно прежними. И это – неопровержимое, полное доказательство того, что хлоропласты и в самом деле когда-то имели прокариотического предка. Возможно, это был организм, от которого произошли также современные цианобактерии.
Развитие хлоропласта из пропластиды
«Взрослый» органоид развивается из пропластиды. Это маленькая, полностью бесцветная органелла, имеющая всего несколько микрон в поперечнике. Она окружена плотной двуслойной мембраной, которая содержит кольцевую ДНК, специфическую для хлоропласта. Внутренней мембранной системы эти «предки» органоидов не имеют. Из-за предельно малых размеров их изучение крайне затруднено, а потому данных об их развитии чрезвычайно мало.
Известно, что несколько таких протопластид имеется в ядре каждой яйцеклетки животных и растений. В ходе развития зародыша они делятся и передаются другим клеткам. Это легко проверить: генетические признаки, которые так или иначе связаны с пластидами, передаются только по материнской линии.
Внутренняя мембрана протопластиды за время развития выпячивается внутрь органоида. Из этих структур вырастают мембраны тилакоидов, которые отвечают за образование гран и ламелл стромы органоида. В полной темноте протопастида начинает преобразовываться в предшественник хлоропласта (этиопласта). Этот первичный органоид характерен тем, что внутри него располагается довольно сложная кристаллическая структура. Как только на лист растения попадет свет, она полностью разрушается. После этого происходит образование «традиционной» внутренней структуры хлоропласта, которая образована как раз-таки тилакоидами и ламеллами.







